
本期介绍的“细胞焦亡”,有可能给肿瘤和免疫治疗领域带来一个全新的机会。靶向细胞焦亡的药物属于全新生物学机制的“first in category”药物。如何成功开发,考验的不光是科学发现的质量,还有科学家、药物开发者、临床医生之间的密切合作。
撰文 |王立铭
众所周知,细胞是地球生命的基本组成单元。人类这样的多细胞生物,每个个体都由数十万亿个细胞按照复杂的时空组织逻辑构建而来,这些细胞的使命就是竭尽全力保证我们的出生、成长、和繁殖。为了实现这些伟大的目标,细胞生命本身是随时可以牺牲和替代的;在很多场合,身体细胞甚至还会主动开启死亡程序来协助人体更好地生存和繁殖。
本期报告要重点介绍的“细胞焦亡”,也是人体细胞内生的一种死亡程序。这种特殊的细胞死亡程序在1990年代逐渐为人所知。研究者们发现,人体免疫细胞在被志贺杆菌、沙门氏菌等病原菌入侵后,会引发细胞死亡。这种情境下出现的细胞死亡具有鲜明的形态特征:进入死亡程序的细胞,细胞体积快速涨大,细胞膜上出现穿孔和鼓包,直至细胞的完整性彻底被破坏,细胞内容物大量泄露,引发周围组织的炎症反应。显然这是一种非常“脏”和“暴烈”的死亡方式,其英文名称(pyroptosis)也隐含了这层意思:希腊语词根pyro-指“火焰”,而-ptosis则指“跌落”。
细胞焦亡和人体免疫系统对病原体的防御有密切关系。人体免疫系统的很多细胞,如巨噬细胞、中心粒细胞、树突状细胞、B细胞和T细胞等,在正常状态下可以直接识别和杀伤病原微生物。但如果它们自己也被病原微生物感染且无力清除时,可能会选择焦亡这类暴烈的死亡方式。在和病原体同归于尽的同时,也向周围环境释放了明确的危险信号,进一步吸引免疫系统的火力,召集和激活更多的免疫细胞,帮助机体更好地清除病原体威胁。
在过去20年时间内,人们也开始逐渐理解细胞焦亡具体是如何发生的。这些信息除了让人们得以理解细胞焦亡的全貌,也为利用细胞焦亡过程开发肿瘤和免疫相关药物奠定了基础。
这里我们简要的回顾和总结一下细胞焦亡研究路程中的关键性节点。
在2011年,美国基因泰克公司的Vishva Dixit实验室发现,多种革兰氏阴性菌(如大肠杆菌、啮齿类柠檬酸杆菌、霍乱弧菌)的感染,会诱导小鼠的巨噬细胞释放大量促进炎症反应的细胞因子如白介素1b和白介素18等,也会诱导巨噬细胞的死亡[1]。在2013年和2014年,Dixit实验室和邵峰实验室又先后发现,被革兰氏阴性菌感染后,革兰氏阴性菌表面的脂多糖分子(LPS)可以结合和激活巨噬细胞内的一个特殊蛋白酶(caspase-4/11),从而引发巨噬细胞的焦亡[2][3]。这是人们第一次开始理解细胞焦亡的具体机制。
而细胞焦亡本身又是如何发生的呢?
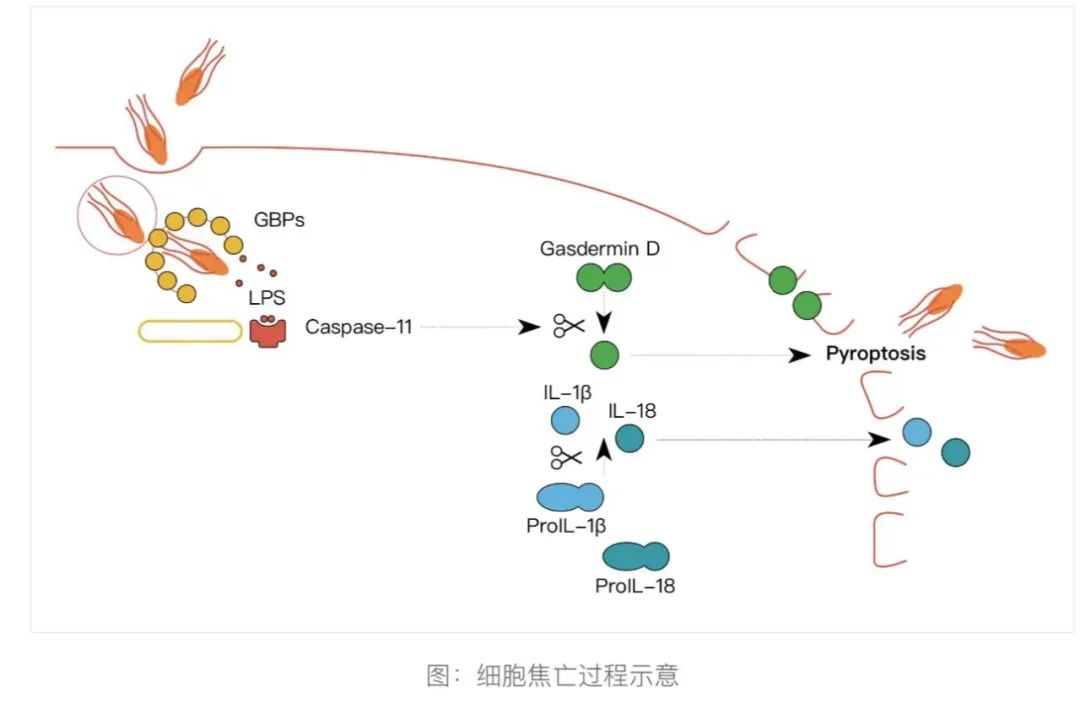
2015年,Dixit实验室和邵峰实验室利用截然不同的实验手段,证明细胞焦亡是被激活的caspase蛋白酶切割一个名叫Gasdermin D的蛋白质所引发的[4][5]。具体而言,Gasdermin D是一个接近500个氨基酸长度的蛋白质,在中间部位有一段序列可以被caspase蛋白酶切割。在未切割状态下,Gasdermin D的前半部分和后半部分可以相互结合,进入一种功能深度抑制的结构。一旦被切割开后,Gasdermin D的前半部分蛋白就被释放出来,直接推动焦亡的发生。需要顺便说明的是,在2015年之前,Gasdermin蛋白的功能几乎是完全的空白,Dixit实验室是通过小鼠突变体筛选的方法寻找对LPS不敏感的基因缺陷找到了Gasdermin D,而邵峰实验室则是通过当时刚刚发明的CRISPR/cas9筛选的方法在细胞系中找到了Gasdermin D。
在2016年,被切割的Gasdermin D如何导致焦亡也有了更进一步的信息。数个研究组(包括邵峰实验室和Dixit实验室,还有哈佛大学医学院的吴皓实验室和Judy Lieberman实验室等)在同一时间发现,Gasdermin D在被蛋白酶切割后,其前半段能够插入细胞膜,大约27个Gasdermin D单体能够组装形成一个直径18纳米的巨型孔洞,正是这个孔洞使得了细胞外水分涌入细胞,细胞膜膨大,细胞内容物泄露,最终导致细胞的暴裂死亡[6][7]。在2018年,吴皓实验室还详细解析了Gasdermin聚集成环形成细胞孔洞的结构信息[8]。
至此,细胞焦亡发生的完整脉络得到了充分的说明,在此历程中,邵峰实验室、Vishva Dixit实验室、Judy Lieberman实验室和吴皓实验室作出了重要的奠基性工作。其中特别是邵峰和Vishva Dixit实验室找到了焦亡的具体执行者,即在被切割后能够组装成环、引起细胞破孔的Gasdermin蛋白。这些发现为我们深入理解细胞焦亡在免疫和肿瘤中的具体生物学功能,以及响应的药物开发铺平了道路。
我们可以粗略的将细胞焦亡的过程分为三个步骤:
启动阶段:病原体入侵细胞,暴露出其特征识别信号(如革兰氏阴性菌表面的脂多糖分子);
准备阶段:细胞内防御机制识别出上述信号,并开始准备焦亡所需的元件(如脂多糖分子被caspase蛋白酶所识别,进一步切割Gasdermin D蛋白);
完成阶段:细胞膜结构破裂,导致细胞焦亡(如被切割后的Gasdermin D蛋白插入细胞膜,形成孔洞)。
值得特别指出的是,在上述“主干”之外,研究者们也陆续发现细胞焦亡的每个步骤也有不同的分支,让我们对细胞焦亡在免疫反应中的角色有了更深刻的理解。
例如,除了革兰氏阴性菌的脂多糖之外,病原体分泌的毒素(例如炭疽毒素和志贺毒素)、细胞内游离的RNA和双链DNA(往往来自病毒感染)、细菌的特征性结构(例如长长的鞭毛和状如注射器的3型/5型分泌系统)、病原体代谢产物(例如合成脂多糖过程的中间产物ADP-庚糖)等,都能通过不同的细胞内检测机制被识别,启动炎症反应和细胞焦亡[9][10][11][12]。
同时,在焦亡的准备阶段,除了上文介绍过的caspase-4/11蛋白酶,其他类型的caspase蛋白酶(例如caspase-1/3/8),在特定条件下也能通过切割Gasdermin,引起炎症反应。还有研究提示在某些条件下Gasdermin的切割甚至可以完全摆脱对caspase的依赖。例如,在肿瘤免疫微环境中,免疫细胞可以通过直接释放颗粒酶进入肿瘤细胞,切割Gasdermin从而诱导细胞焦亡[13]。换句话说,焦亡的第二阶段主要服务于Gasdermin蛋白的切割和激活,具体的操作途径则是比较灵活的。
还有,在上述这些多样化的启动和准备分支之后,细胞也能引发不同类型的免疫反应。例如细胞能通过释放白介素1b和白介素18等方式吸引和激活周围的免疫细胞,促进机体的炎症反应,以更好地清除病原体入侵。当然,这里的重点是我们重点介绍的细胞焦亡,而细胞焦亡的最终“出口”都要依靠Gasdermin蛋白的组装成环。除了上面描述的Gasdermin D之外,人类基因组中还有四个Gasdermin家族成员(A/B/C/E)都表现出了类似的引发焦亡的功能[14]。
根据如上的分析,我们其实可以在人体天然免疫这个更大的框架下理解焦亡。所谓天然免疫,指的是不依赖后天经验和训练,直接识别入侵异源物质的某些固有特征,并对其进行清除和杀伤的机制。在这个框架下,焦亡可以看成是天然免疫系统的“兜底”措施。
图:细胞焦亡和机体天然免疫系统(https://www.nature.com/articles/s41423-021-00740-6)
当人体的天然免疫系统,例如巨噬细胞,识别到了人体中正常条件下不会出现、往往由入侵病原体携带的某些特征(诸如革兰氏阴性菌的脂多糖外壳),就会启动一系列免疫反应对其进行杀伤。如果上述防御机制没有起到期望中的效果,巨噬细胞就可能启动自身的焦亡,用一种更加暴烈的方式杀伤内部的病原菌,并将能够引发免疫反应的信号更强烈地释放出去,以激发更大范围的免疫反应。
如上所述,我们可以把细胞焦亡看成人体应对病原体入侵的免疫防御机制的一部分。和人体其他类型的免疫机制类似,焦亡也需要被小心地维持在一个合理的强度。过强(“不该死的死了”),可能会产生过度的免疫反应,引发慢性炎症和自身免疫性疾病;而过弱(“该死的没死”),则可能促进癌细胞的繁殖复制,或者影响了肿瘤治疗方案的效果。
这当然不是焦亡特有的属性。在过去二三十时间里,围绕免疫系统的过度活跃或者抑制,人们阐明了许多疾病的发病机理,也开发出了相应的药物。
针对免疫系统的过度活跃,一个经典案例是1960-70年代发现的肿瘤坏死因子TNF-α。和上面介绍的焦亡过程类似,TNF-α也是机体抵御病原体入侵的重要防御手段。在诸如细菌脂多糖等危险信号的刺激下,TNF-α主要由巨噬细胞等免疫细胞大量产生和分泌,吸引和激活各种免疫细胞,引发机体炎症反应,以消灭入侵病原体。但TNF-α的异常增高也能导致一系列慢性炎症和自身免疫疾病,因此靶向TNF-α也成为治疗这些疾病的重要途径。在医疗市场上,多个抑制TNF-α的药物,诸如阿达木单抗(Adalimumab)、英夫利昔单抗(Infliximab)、依那西普(Etanercept)等,都是横跨类风湿性关节炎、强直性脊柱炎、银屑病性关节炎、克罗恩病、溃疡性结肠炎等多种炎症适应症的重磅药物[15]。
后者的经典应用则更广为人知。1990年代,研究者们先后发现了CTLA4和PD-1两个在免疫细胞中表达的细胞表面蛋白,且都介导了对免疫机能的抑制作用。敲除CTLA4或PD-1的小鼠都表现出了全身性的自身免疫症状。后续研究更是进一步发现,肿瘤细胞可以通过高表达PD-1的配体PD-L1的方式抑制免疫机能,逃逸免疫系统的识别和杀伤,从而促进自身的生存和繁殖。这一系列研究最终指向了引发肿瘤治疗领域革命的癌症免疫疗法。靶向PD-1/PD-L1通路的药物纳武单抗(Nivolumab)、帕博丽珠单抗(Pembrolizumab),靶向CTLA通路的伊匹单抗(lpilimumab)等,也成为了适应症覆盖十几种不同肿瘤类型的重磅药物[16]。
延续类似的逻辑,细胞焦亡的异常激活或者抑制,或者更广泛的天然免疫系统的异常激活和抑制,是否有可能和人类疾病有直接关联,从而可以用于指导药物的开发呢?
当然如此。我们仍然先讨论细胞焦亡过程的异常激活。研究者们已经发现,天然免疫和焦亡通路的蛋白,如果出现功能增强的基因变异,确实可能会导致严重的自身免疫疾病。例如Pyrin的基因变异会导致家族性地中海热,发病过程中也伴随着Gasdermin D介导的细胞焦亡[17];ALPK1的基因变异会导致罕见的ROSAH综合征[18],等等。在更为常见的自身免疫病,例如类风湿性关节炎、溃疡性结肠炎中,也观察到了细胞焦亡过程的异常激活[19][20][21]。还有,病原微生物入侵人体如果长时间得不到有效清除,人体的天然免疫反应和焦亡过程会过度激活,引起致命的脓毒血症[22]。那么,设计药物反其道而行之,抑制焦亡的过度激活,也许就能够治疗这些疾病。
细胞焦亡的异常抑制也和人类疾病有关。在不少肿瘤类型中,焦亡通路的基因,特别是Gasdermin家族,处于深度抑制状态[23]。这本身很容易理解,因为诱导细胞死亡,包括凋亡和焦亡,本就是人体免疫系统消灭异常细胞的手段之一。癌细胞自然需要通过逃逸这些控制手段来保护自身的生存和繁殖。而相应的,2020年来自邵峰实验室和Judy Lieberman实验室的两项研究也分别证明,免疫细胞也可以通过一个非常特别的机制绕过上述逃逸机制杀伤肿瘤细胞。具体来说,免疫细胞可以释放一类叫做颗粒素(Granzyme)的蛋白酶,借由细胞之间的穿孔素通道进入癌细胞,直接切割并激活Gasdermin蛋白,诱导肿瘤细胞的焦亡[24][25]。
按照此逻辑推演,激活肿瘤细胞内被深度抑制的焦亡通路,是一条很有前途的肿瘤药物开发路径。事实上,2020年人们也发现,如果将Gasdermin蛋白特异性的导入肿瘤细胞,确实观察到了显著的肿瘤杀伤效果[26]。特别值得注意的是,肿瘤细胞焦亡可能不仅仅是杀死了这枚肿瘤细胞自己。我们在前文反复提到,细胞焦亡这种暴烈的细胞死亡过程,能够最大限度的刺激机体免疫反应。一枚肿瘤细胞的焦亡,可能发挥所谓的“旁观者效应”,吸引和激活更多的免疫细胞来到肿瘤位置,杀伤周围更多的肿瘤细胞。作为旁证,2017年的一项研究也证明,化疗药物的肿瘤杀伤作用,也可能是通过诱导焦亡、激活病灶部位的免疫反应来实现的[27]。
值得一提的是,不同的Gasdermin家族成员体现出了不同的组织特异性,这一点既反映了它们不同的生理功能,也为后续药物开发给出了重要的提示。
例如,Gasdermin D的表达范围最为广泛,因此可以用来治疗全身性的炎症疾病,例如病原菌感染引起的脓毒血症[28];Gasdermin B的基因变异和异常表达和哮喘、溃疡性结肠炎等慢性炎症的发病有关,因此对它的抑制可能治疗这些疾病[29];而Gasdermin B/E在某些肿瘤类型中高表达(但功能被深度抑制),因此如果设计药物激活它们则可能成为有效的癌症药物。
细胞焦亡作为药物靶点:展望和挑战
根据上面的讨论,细胞焦亡作为肿瘤和免疫相关疾病的全新靶点,在科学上已经有了相当充分的证据。简单总结的话,抑制细胞焦亡通路可能治疗和人体免疫活动过强有关的疾病,包括各种慢性炎症、自身免疫疾病、细菌感染引发的脓毒血症等;而激活细胞焦亡通路则可能杀伤癌细胞,治疗各种癌症,或者提升其他癌症药物的杀伤效率。
当然,这只是一个较为简化的总结。真实的疾病场景和药物开发过程显然要复杂得多。就Gasdermin蛋白家族来说,其促进焦亡的机制并非是简单的酶学反应,而是是被切割后大量单体组装成环、形成细胞膜上的孔洞,因此如何筛选和设计药物阻止Gasdermin的切割和组装,本身就是很有挑战性的任务。同时因为Gasdermin家族有五个成员都可以引起焦亡,可能我们还需要区分不同成员在不同免疫疾病中的作用,设计有靶点专一性的药物。
更重要的是,上面我们讨论的研究固然主要在免疫细胞(特别是天然免疫的核心成员巨噬细胞)中完成,但焦亡通路却是普遍存在于人体细胞中的固有机制。因此如果不加选择地抑制或者激活焦亡通路,可能会引发全身性的毒副作用。因此,在这类药物用于疾病治疗时,可能也需要审慎地对适应症和适用人群进行选择。
这些问题是所有真正原创性的新药上市之前都必须面临和克服的问题。对于靶向全新生物学机制(而非仅仅是全新生物学靶点)的“first in category”药物来说更是如此。
就拿我们刚刚讨论过的肿瘤免疫药物为例,事实上,即便人们已经在实验室研究中理解了CTLA4和PD-1/PD-L1信号确实能够抑制免疫细胞的功能,特别是肿瘤细胞还能够通过PD-L1信号逃逸免疫系统的识别和杀伤,当这些药物进入临床试验后,仍然还需要一个漫长的探索过程去挖掘真正的适应症和适用人群。
靶向CTLA4的伊匹单抗因副作用较大,最终仅在黑色素瘤这一个适应症上获得批准。而靶向PD-1的纳武单抗和帕博丽珠单抗虽然目前获批了多个癌种的适应症,但其临床探索过程也可谓一波三折。主导纳武单抗临床试验的美国BMS公司在一期临床试验中纳入了多个实体瘤类别的受试者,并看到了较为积极的疗效[30],BMS公司一鼓作气推动了更大规模的多瘤种临床试验,但却遭遇了数个意料之外的失败,2016年该药物在非小细胞肺癌中的惨烈失败更是动摇了整个市场对癌症免疫药物的信心[31]。所幸在2015-2017年,约翰霍普金斯大学的Bert Vogelstein实验室证明,存在DNA错配修复障碍、从而带有高突变载荷的肿瘤细胞更容易对PD-1药物产生响应[32][33],才为这类药物找到了真正准确的适用人群。
近年来,在天然免疫领域也有一个类似的明星分子可供参照,就是NLRP3分子。这个分子广泛参与了免疫细胞对多种病原体特征信号(如ATP和双链RNA)、以及人体内异常生理信号(如饱和脂肪酸和尿酸钠结晶)的识别,也能通过激活caspase诱导包括焦亡在内的炎症反应[34]。根据我们上面的讨论,靶向NLRP3分子也可能成为治疗各种慢性炎症和自身免疫疾病的新方式。近年来也有多家公司致力于开发靶向NLRP3的药物,其中不乏资本市场的宠儿,例如在2019年被诺华以15亿美金总价收购的IFM Therapeutics公司,以及2018年上市、市值约15亿美元的Ventyx Biosciences公司。但即便如此,靶向NLRP3的药物目前仍未能明确其适应症和适用人群,估计还需要多个大规模临床试验的摸索才能尘埃落定。
从本质上说,这个探索过程其实是基础科学研究向临床应用转化的必经之路。在实验室里,科学家们日常处理的是变量单一、参数可控的“完美”实验条件,相对容易获得干净和纯粹的研究结论。但一旦进入人体真实应用,特别是诸如慢性炎症和肿瘤这样动态变化、成因复杂、千人千面的疾病,需要考虑的干扰因素骤然变多,而药物本身的影响也更为复杂广泛。如何完成这中间的转换,考验的不光是科学发现的质量,还有科学家、药物开发者、临床医生之间的密切合作。
和此目标相一致的,2020年邵峰也主导创立了炎明生物,旨在开发靶向Gasdermin和ALPK1蛋白的全新药物。根据公开信息,炎明生物已经开发了靶向ALPK1蛋白的小分子激活剂,以及靶向Gasdermin的小分子抑制剂。我当然也非常期待,邵峰实验室积累的科学发现,能够以炎明生物为载体,完成一个万众期待的证明:在中国的土地上,不光能完成高度原创性的生物医学发现,还能通过科学家、药物开发者、临床医生的紧密配合,完成原创性科学发现到“first in category”药物开发的顺利转化,惠泽全世界的疾病患者。
 参考文献:(上下滑动可浏览)[1] Kayagaki, N., Warming, S., Lamkanfi, M., Vande Walle, L., Louie, S., Dong, J., Newton, K., Qu, Y., Liu, J., Heldens, S., Zhang, J., Lee, W. P., Roose-Girma, M., & Dixit, V. M. (2011). Non-canonical inflammasome activation targets caspase-11. Nature, 479(7371), 117–121.[2] Kayagaki, N., Wong, M. T., Stowe, I. B., Ramani, S. R., Gonzalez, L. C., Akashi-Takamura, S., Miyake, K., Zhang, J., Lee, W. P., Muszyński, A., Forsberg, L. S., Carlson, R. W., & Dixit, V. M. (2013). Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science (New York, N.Y.), 341(6151), 1246–1249. [3]Shi, J., Zhao, Y., Wang, Y., Gao, W., Ding, J., Li, P., Hu, L., & Shao, F. (2014). Inflammatory caspases are innate immune receptors for intracellular LPS. Nature, 514(7521), 187–192. [4] Kayagaki, N., Stowe, I. B., Lee, B. L., O'Rourke, K., Anderson, K., Warming, S., Cuellar, T., Haley, B., Roose-Girma, M., Phung, Q. T., Liu, P. S., Lill, J. R., Li, H., Wu, J., Kummerfeld, S., Zhang, J., Lee, W. P., Snipas, S. J., Salvesen, G. S., Morris, L. X., … Dixit, V. M. (2015). Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature, 526(7575), 666–671. [5] Shi, J., Zhao, Y., Wang, K., Shi, X., Wang, Y., Huang, H., Zhuang, Y., Cai, T., Wang, F., & Shao, F. (2015). Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature, 526(7575), 660–665. [6] Ding, J., Wang, K., Liu, W., She, Y., Sun, Q., Shi, J., Sun, H., Wang, D. C., & Shao, F. (2016). Pore-forming activity and structural autoinhibition of the gasdermin family. Nature, 535(7610), 111–116. [7] Liu, X., Zhang, Z., Ruan, J., Pan, Y., Magupalli, V. G., Wu, H., & Lieberman, J. (2016). Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature, 535(7610), 153–158. [8] Ruan, J., Xia, S., Liu, X., Lieberman, J., & Wu, H. (2018). Cryo-EM structure of the gasdermin A3 membrane pore. Nature (London), 557(7703), 62–67. [9] Yu, P., Zhang, X., Liu, N., Tang, L., Peng, C., & Chen, X. (2021). Pyroptosis: mechanisms and diseases. Signal Transduction and Targeted Therapy, 6(1), 128–128. [10] Zhao, Y., Yang, J., Shi, J., Gong, Y. N., Lu, Q., Xu, H., Liu, L., & Shao, F. (2011). The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nature, 477(7366), 596–600. [11] Xu, H., Yang, J., Gao, W., Li, L., Li, P., Zhang, L., Gong, Y. N., Peng, X., Xi, J. J., Chen, S., Wang, F., & Shao, F. (2014). Innate immune sensing of bacterial modifications of Rho GTPases by the Pyrin inflammasome. Nature, 513(7517), 237–241. [12] Zhou, P., She, Y., Dong, N., Li, P., He, H., Borio, A., Wu, Q., Lu, S., Ding, X., Cao, Y., Xu, Y., Gao, W., Dong, M., Ding, J., Wang, D. C., Zamyatina, A., & Shao, F. (2018). Alpha-kinase 1 is a cytosolic innate immune receptor for bacterial ADP-heptose. Nature, 561(7721), 122–126. [13] Zhou, Z., He, H., Wang, K., Shi, X., Wang, Y., Su, Y., Wang, Y., Li, D., Liu, W., Zhang, Y., Shen, L., Han, W., Shen, L., Ding, J., & Shao, F. (2020). Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science (New York, N.Y.), 368(6494), eaaz7548.[14] De Schutter, E., Roelandt, R., Riquet, F. B., Van Camp, G., Wullaert, A., & Vandenabeele, P. (2021). Punching Holes in Cellular Membranes: Biology and Evolution of Gasdermins. Trends in Cell Biology, 31(6), 500–513. [15] Li, P., Zheng, Y., & Chen, X. (2017). Drugs for autoimmune inflammatory diseases: From small molecule compounds to anti-TNF biologics. Frontiers in Pharmacology, 8, 460–460. [16] Wojtukiewicz MZ, Rek MM, Karpowicz K, Górska M, Polityńska B, Wojtukiewicz AM, Moniuszko M, Radziwon P, Tucker SC, Honn KV. Inhibitors of immune checkpoints-PD-1, PD-L1, CTLA-4-new opportunities for cancer patients and a new challenge for internists and general practitioners. Cancer Metastasis Rev. 2021 Sep;40(3):949-982. doi: 10.1007/s10555-021-09976-0. Epub 2021 Jul 8. PMID: 34236546; PMCID: PMC8556173.[17] Kanneganti, A., Malireddi, R., Saavedra, P., Vande Walle, L., Van Gorp, H., Kambara, H., Tillman, H., Vogel, P., Luo, H. R., Xavier, R. J., Chi, H., & Lamkanfi, M. (2018). GSDMD is critical for autoinflammatory pathology in a mouse model of Familial Mediterranean Fever. The Journal of experimental medicine, 215(6), 1519–1529. [18] Williams, L. B., Javed, A., Sabri, A., Morgan, D. J., Huff, C. D., Grigg, J. R., Heng, X. T., Khng, A. J., Hollink, I., Morrison, M. A., Owen, L. A., Anderson, K., Kinard, K., Greenlees, R., Novacic, D., Nida Sen, H., Zein, W. M., Rodgers, G. M., Vitale, A. T., Haider, N. B., … DeAngelis, M. M. (2019). ALPK1 missense pathogenic variant in five families leads to ROSAH syndrome, an ocular multisystem autosomal dominant disorder. Genetics in medicine : official journal of the American College of Medical Genetics, 21(9), 2103–2115. [19] Gu, L., Sun, Y., Wu, T., Chen, G., Tang, X., Zhao, L., He, L., Hu, Z., Sun, L., Pan, F., Yin, Z., & Guo, Z. (2022). A novel mechanism for macrophage pyroptosis in rheumatoid arthritis induced by Pol β deficiency. Cell Death & Disease, 13(7), 583–583. [20] Tan, G., Huang, C., Chen, J., Chen, B., & Zhi, F. (2021). Gasdermin-E-mediated pyroptosis participates in the pathogenesis of Crohn's disease by promoting intestinal inflammation. Cell reports, 35(11), 109265. [21] Das, S., Miller, M., Beppu, A. K., Mueller, J., McGeough, M. D., Vuong, C., Karta, M. R., Rosenthal, P., Chouiali, F., Doherty, T. A., Kurten, R. C., Hamid, Q., Hoffman, H. M., & Broide, D. H. (2016). GSDMB induces an asthma phenotype characterized by increased airway responsiveness and remodeling without lung inflammation. Proceedings of the National Academy of Sciences of the United States of America, 113(46), 13132–13137. [22] Zheng X, Chen W, Gong F, Chen Y, Chen E. The Role and Mechanism of Pyroptosis and Potential Therapeutic Targets in Sepsis: A Review. Front Immunol. 2021 Jul 7;12:711939. doi: 10.3389/fimmu.2021.711939. PMID: 34305952; PMCID: PMC8293747.[23] Xia, X., Wang, X., Cheng, Z., Qin, W., Lei, L., Jiang, J., & Hu, J. (2019). The role of pyroptosis in cancer: pro-cancer or pro-“host”? Cell Death & Disease, 10(9), 650–13. [24] Zhou, Z., He, H., Wang, K., Shi, X., Wang, Y., Su, Y., Wang, Y., Li, D., Liu, W., Zhang, Y., Shen, L., Han, W., Shen, L., Ding, J., & Shao, F. (2020). Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science (New York, N.Y.), 368(6494), eaaz7548.[25] Zhang, Z., Zhang, Y., Xia, S., Kong, Q., Li, S., Liu, X., Junqueira, C., Meza-Sosa, K. F., Mok, T., Ansara, J., Sengupta, S., Yao, Y., Wu, H., & Lieberman, J. (2020). Gasdermin E suppresses tumour growth by activating anti-tumour immunity. Nature, 579(7799), 415–420. [26] Wang, Q., Wang, Y., Ding, J., Wang, C., Zhou, X., Gao, W., Huang, H., Shao, F., & Liu, Z. (2020). A bioorthogonal system reveals antitumour immune function of pyroptosis. Nature, 579(7799), 421–426. [27] Wang, Y., Gao, W., Shi, X., Ding, J., Liu, W., He, H., Wang, K., & Shao, F. (2017). Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature (London), 547(7661), 99–103. [28] Hu, J. J., Liu, X., Xia, S., Zhang, Z., Zhang, Y., Zhao, J., Ruan, J., Luo, X., Lou, X., Bai, Y., Wang, J., Hollingsworth, L. R., Magupalli, V. G., Zhao, L., Luo, H. R., Kim, J., Lieberman, J., & Wu, H. (2020). FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation. Nature Immunology, 21(7), 736–745. [29] Chao, K. L., Kulakova, L., & Herzberg, O. (2017). Gene polymorphism linked to increased asthma and IBD risk alters gasdermin-B structure, a sulfatide and phosphoinositide binding protein. Proceedings of the National Academy of Sciences - PNAS, 114(7), E1128–E1137. [30] Brahmer, J. R., Drake, C. G., Wollner, I., Powderly, J. D., Picus, J., Sharfman, W. H., Stankevich, E., Pons, A., Salay, T. M., McMiller, T. L., Gilson, M. M., Wang, C., Selby, M., Taube, J. M., Anders, R., Chen, L., Korman, A. J., Pardoll, D. M., Lowy, I., & Topalian, S. L. (2010). Phase I study of single-agent anti-programmed death-1 (MDX-1106) in refractory solid tumors: safety, clinical activity, pharmacodynamics, and immunologic correlates. Journal of clinical oncology : official journal of the American Society of Clinical Oncology, 28(19), 3167–3175. [31] Carbone, D. P., Reck, M., Paz-Ares, L., Creelan, B., Horn, L., Steins, M., Felip, E., van den Heuvel, M. M., Ciuleanu, T.-E., Badin, F., Ready, N., Hiltermann, T. J. N., Nair, S., Juergens, R., Peters, S., Minenza, E., Wrangle, J. M., Rodriguez-Abreu, D., Borghaei, H., … Socinski, M. A. (2017). First-Line Nivolumab in Stage IV or Recurrent Non–Small-Cell Lung Cancer. The New England Journal of Medicine, 376(25), 2415–2426. [32] Le, D. T., Uram, J. N., Wang, H., Bartlett, B. R., Kemberling, H., Eyring, A. D., Skora, A. D., Luber, B. S., Azad, N. S., Laheru, D., Biedrzycki, B., Donehower, R. C., Zaheer, A., Fisher, G. A., Crocenzi, T. S., Lee, J. J., Duffy, S. M., Goldberg, R. M., de la Chapelle, A., Koshiji, M., … Diaz, L. A., Jr (2015). PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. The New England journal of medicine, 372(26), 2509–2520. [33] Le, D. T., Durham, J. N., Smith, K. N., Wang, H., Bartlett, B. R., Aulakh, L. K., Lu, S., Kemberling, H., Wilt, C., Luber, B. S., Wong, F., Azad, N. S., Rucki, A. A., Laheru, D., Donehower, R., Zaheer, A., Fisher, G. A., Crocenzi, T. S., Lee, J. J., Greten, T. F., … Diaz, L. A., Jr (2017). Mismatch repair deficiency predicts response of solid tumors to PD-1 blockade. Science (New York, N.Y.), 357(6349), 409–413.[34] Swanson, K. V., Deng, M., & Ting, J. P.-Y. (2019). The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nature Reviews. Immunology, 19(8), 477–489.
参考文献:(上下滑动可浏览)[1] Kayagaki, N., Warming, S., Lamkanfi, M., Vande Walle, L., Louie, S., Dong, J., Newton, K., Qu, Y., Liu, J., Heldens, S., Zhang, J., Lee, W. P., Roose-Girma, M., & Dixit, V. M. (2011). Non-canonical inflammasome activation targets caspase-11. Nature, 479(7371), 117–121.[2] Kayagaki, N., Wong, M. T., Stowe, I. B., Ramani, S. R., Gonzalez, L. C., Akashi-Takamura, S., Miyake, K., Zhang, J., Lee, W. P., Muszyński, A., Forsberg, L. S., Carlson, R. W., & Dixit, V. M. (2013). Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science (New York, N.Y.), 341(6151), 1246–1249. [3]Shi, J., Zhao, Y., Wang, Y., Gao, W., Ding, J., Li, P., Hu, L., & Shao, F. (2014). Inflammatory caspases are innate immune receptors for intracellular LPS. Nature, 514(7521), 187–192. [4] Kayagaki, N., Stowe, I. B., Lee, B. L., O'Rourke, K., Anderson, K., Warming, S., Cuellar, T., Haley, B., Roose-Girma, M., Phung, Q. T., Liu, P. S., Lill, J. R., Li, H., Wu, J., Kummerfeld, S., Zhang, J., Lee, W. P., Snipas, S. J., Salvesen, G. S., Morris, L. X., … Dixit, V. M. (2015). Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature, 526(7575), 666–671. [5] Shi, J., Zhao, Y., Wang, K., Shi, X., Wang, Y., Huang, H., Zhuang, Y., Cai, T., Wang, F., & Shao, F. (2015). Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature, 526(7575), 660–665. [6] Ding, J., Wang, K., Liu, W., She, Y., Sun, Q., Shi, J., Sun, H., Wang, D. C., & Shao, F. (2016). Pore-forming activity and structural autoinhibition of the gasdermin family. Nature, 535(7610), 111–116. [7] Liu, X., Zhang, Z., Ruan, J., Pan, Y., Magupalli, V. G., Wu, H., & Lieberman, J. (2016). Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature, 535(7610), 153–158. [8] Ruan, J., Xia, S., Liu, X., Lieberman, J., & Wu, H. (2018). Cryo-EM structure of the gasdermin A3 membrane pore. Nature (London), 557(7703), 62–67. [9] Yu, P., Zhang, X., Liu, N., Tang, L., Peng, C., & Chen, X. (2021). Pyroptosis: mechanisms and diseases. Signal Transduction and Targeted Therapy, 6(1), 128–128. [10] Zhao, Y., Yang, J., Shi, J., Gong, Y. N., Lu, Q., Xu, H., Liu, L., & Shao, F. (2011). The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nature, 477(7366), 596–600. [11] Xu, H., Yang, J., Gao, W., Li, L., Li, P., Zhang, L., Gong, Y. N., Peng, X., Xi, J. J., Chen, S., Wang, F., & Shao, F. (2014). Innate immune sensing of bacterial modifications of Rho GTPases by the Pyrin inflammasome. Nature, 513(7517), 237–241. [12] Zhou, P., She, Y., Dong, N., Li, P., He, H., Borio, A., Wu, Q., Lu, S., Ding, X., Cao, Y., Xu, Y., Gao, W., Dong, M., Ding, J., Wang, D. C., Zamyatina, A., & Shao, F. (2018). Alpha-kinase 1 is a cytosolic innate immune receptor for bacterial ADP-heptose. Nature, 561(7721), 122–126. [13] Zhou, Z., He, H., Wang, K., Shi, X., Wang, Y., Su, Y., Wang, Y., Li, D., Liu, W., Zhang, Y., Shen, L., Han, W., Shen, L., Ding, J., & Shao, F. (2020). Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science (New York, N.Y.), 368(6494), eaaz7548.[14] De Schutter, E., Roelandt, R., Riquet, F. B., Van Camp, G., Wullaert, A., & Vandenabeele, P. (2021). Punching Holes in Cellular Membranes: Biology and Evolution of Gasdermins. Trends in Cell Biology, 31(6), 500–513. [15] Li, P., Zheng, Y., & Chen, X. (2017). Drugs for autoimmune inflammatory diseases: From small molecule compounds to anti-TNF biologics. Frontiers in Pharmacology, 8, 460–460. [16] Wojtukiewicz MZ, Rek MM, Karpowicz K, Górska M, Polityńska B, Wojtukiewicz AM, Moniuszko M, Radziwon P, Tucker SC, Honn KV. Inhibitors of immune checkpoints-PD-1, PD-L1, CTLA-4-new opportunities for cancer patients and a new challenge for internists and general practitioners. Cancer Metastasis Rev. 2021 Sep;40(3):949-982. doi: 10.1007/s10555-021-09976-0. Epub 2021 Jul 8. PMID: 34236546; PMCID: PMC8556173.[17] Kanneganti, A., Malireddi, R., Saavedra, P., Vande Walle, L., Van Gorp, H., Kambara, H., Tillman, H., Vogel, P., Luo, H. R., Xavier, R. J., Chi, H., & Lamkanfi, M. (2018). GSDMD is critical for autoinflammatory pathology in a mouse model of Familial Mediterranean Fever. The Journal of experimental medicine, 215(6), 1519–1529. [18] Williams, L. B., Javed, A., Sabri, A., Morgan, D. J., Huff, C. D., Grigg, J. R., Heng, X. T., Khng, A. J., Hollink, I., Morrison, M. A., Owen, L. A., Anderson, K., Kinard, K., Greenlees, R., Novacic, D., Nida Sen, H., Zein, W. M., Rodgers, G. M., Vitale, A. T., Haider, N. B., … DeAngelis, M. M. (2019). ALPK1 missense pathogenic variant in five families leads to ROSAH syndrome, an ocular multisystem autosomal dominant disorder. Genetics in medicine : official journal of the American College of Medical Genetics, 21(9), 2103–2115. [19] Gu, L., Sun, Y., Wu, T., Chen, G., Tang, X., Zhao, L., He, L., Hu, Z., Sun, L., Pan, F., Yin, Z., & Guo, Z. (2022). A novel mechanism for macrophage pyroptosis in rheumatoid arthritis induced by Pol β deficiency. Cell Death & Disease, 13(7), 583–583. [20] Tan, G., Huang, C., Chen, J., Chen, B., & Zhi, F. (2021). Gasdermin-E-mediated pyroptosis participates in the pathogenesis of Crohn's disease by promoting intestinal inflammation. Cell reports, 35(11), 109265. [21] Das, S., Miller, M., Beppu, A. K., Mueller, J., McGeough, M. D., Vuong, C., Karta, M. R., Rosenthal, P., Chouiali, F., Doherty, T. A., Kurten, R. C., Hamid, Q., Hoffman, H. M., & Broide, D. H. (2016). GSDMB induces an asthma phenotype characterized by increased airway responsiveness and remodeling without lung inflammation. Proceedings of the National Academy of Sciences of the United States of America, 113(46), 13132–13137. [22] Zheng X, Chen W, Gong F, Chen Y, Chen E. The Role and Mechanism of Pyroptosis and Potential Therapeutic Targets in Sepsis: A Review. Front Immunol. 2021 Jul 7;12:711939. doi: 10.3389/fimmu.2021.711939. PMID: 34305952; PMCID: PMC8293747.[23] Xia, X., Wang, X., Cheng, Z., Qin, W., Lei, L., Jiang, J., & Hu, J. (2019). The role of pyroptosis in cancer: pro-cancer or pro-“host”? Cell Death & Disease, 10(9), 650–13. [24] Zhou, Z., He, H., Wang, K., Shi, X., Wang, Y., Su, Y., Wang, Y., Li, D., Liu, W., Zhang, Y., Shen, L., Han, W., Shen, L., Ding, J., & Shao, F. (2020). Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science (New York, N.Y.), 368(6494), eaaz7548.[25] Zhang, Z., Zhang, Y., Xia, S., Kong, Q., Li, S., Liu, X., Junqueira, C., Meza-Sosa, K. F., Mok, T., Ansara, J., Sengupta, S., Yao, Y., Wu, H., & Lieberman, J. (2020). Gasdermin E suppresses tumour growth by activating anti-tumour immunity. Nature, 579(7799), 415–420. [26] Wang, Q., Wang, Y., Ding, J., Wang, C., Zhou, X., Gao, W., Huang, H., Shao, F., & Liu, Z. (2020). A bioorthogonal system reveals antitumour immune function of pyroptosis. Nature, 579(7799), 421–426. [27] Wang, Y., Gao, W., Shi, X., Ding, J., Liu, W., He, H., Wang, K., & Shao, F. (2017). Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature (London), 547(7661), 99–103. [28] Hu, J. J., Liu, X., Xia, S., Zhang, Z., Zhang, Y., Zhao, J., Ruan, J., Luo, X., Lou, X., Bai, Y., Wang, J., Hollingsworth, L. R., Magupalli, V. G., Zhao, L., Luo, H. R., Kim, J., Lieberman, J., & Wu, H. (2020). FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation. Nature Immunology, 21(7), 736–745. [29] Chao, K. L., Kulakova, L., & Herzberg, O. (2017). Gene polymorphism linked to increased asthma and IBD risk alters gasdermin-B structure, a sulfatide and phosphoinositide binding protein. Proceedings of the National Academy of Sciences - PNAS, 114(7), E1128–E1137. [30] Brahmer, J. R., Drake, C. G., Wollner, I., Powderly, J. D., Picus, J., Sharfman, W. H., Stankevich, E., Pons, A., Salay, T. M., McMiller, T. L., Gilson, M. M., Wang, C., Selby, M., Taube, J. M., Anders, R., Chen, L., Korman, A. J., Pardoll, D. M., Lowy, I., & Topalian, S. L. (2010). Phase I study of single-agent anti-programmed death-1 (MDX-1106) in refractory solid tumors: safety, clinical activity, pharmacodynamics, and immunologic correlates. Journal of clinical oncology : official journal of the American Society of Clinical Oncology, 28(19), 3167–3175. [31] Carbone, D. P., Reck, M., Paz-Ares, L., Creelan, B., Horn, L., Steins, M., Felip, E., van den Heuvel, M. M., Ciuleanu, T.-E., Badin, F., Ready, N., Hiltermann, T. J. N., Nair, S., Juergens, R., Peters, S., Minenza, E., Wrangle, J. M., Rodriguez-Abreu, D., Borghaei, H., … Socinski, M. A. (2017). First-Line Nivolumab in Stage IV or Recurrent Non–Small-Cell Lung Cancer. The New England Journal of Medicine, 376(25), 2415–2426. [32] Le, D. T., Uram, J. N., Wang, H., Bartlett, B. R., Kemberling, H., Eyring, A. D., Skora, A. D., Luber, B. S., Azad, N. S., Laheru, D., Biedrzycki, B., Donehower, R. C., Zaheer, A., Fisher, G. A., Crocenzi, T. S., Lee, J. J., Duffy, S. M., Goldberg, R. M., de la Chapelle, A., Koshiji, M., … Diaz, L. A., Jr (2015). PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. The New England journal of medicine, 372(26), 2509–2520. [33] Le, D. T., Durham, J. N., Smith, K. N., Wang, H., Bartlett, B. R., Aulakh, L. K., Lu, S., Kemberling, H., Wilt, C., Luber, B. S., Wong, F., Azad, N. S., Rucki, A. A., Laheru, D., Donehower, R., Zaheer, A., Fisher, G. A., Crocenzi, T. S., Lee, J. J., Greten, T. F., … Diaz, L. A., Jr (2017). Mismatch repair deficiency predicts response of solid tumors to PD-1 blockade. Science (New York, N.Y.), 357(6349), 409–413.[34] Swanson, K. V., Deng, M., & Ting, J. P.-Y. (2019). The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nature Reviews. Immunology, 19(8), 477–489.